Research
We are a group of ecologists and evolutionary biologists who work to describe patterns of biological diversity, identify the evolutionary processes responsible for these patterns, and produce knowledge that communities and managers can use for conservation. Work in the lab typically focuses on birds or mammals in Western North America or the tropical Pacific. Though our projects are diverse, they are united by a combination of fieldwork, genomics, and data science. We also value natural history museums and natural history, reproducibility and best practices in scientific programming, and writing clearly about science for diverse audiences. Major research themes and active projects are described below. PDFs of all papers and brief descriptions of their contents can be found on the Publications page.
Conservation and Biodiversity Genomics
Genomes are a rich source of information on the history, reproductive behavior, and fitness of threatened and endangered species. These data can be used by managers and other stakeholders to identify populations of conservation significance, assess their vulnerability, and plan conservation interventions. Conservation genomics is a major research focus for the Linck Lab, typically conducted in collaboration with land management agencies in the jurisdictional United States. Previous work in this area includes assembly of a highly contiguous reference genome of Clark’s Nutcracker (Innes et al. 2026; with Peter Innes and Matt Carling) and assessment of geographic and political biases in published reference genomes (Linck & Cadena 2025; with Daniel Cadena).
Current projects:
- Conservation genomics of Guam’s sole surviving native forest bird, the Såli (led by M.S. student Jason Gregg and in collaboration with Haldre Rogers and others)
- Genetic monitoring of a reintroduced swift fox population on Fort Belnap Indian Reservation (led by M.S. student Owen Kanter and in collaboration with Justine Becker, Jesse Boulerice, and Nuch Songsasen, and others; funded by the Smithsonian’s Great Plains Science Program)
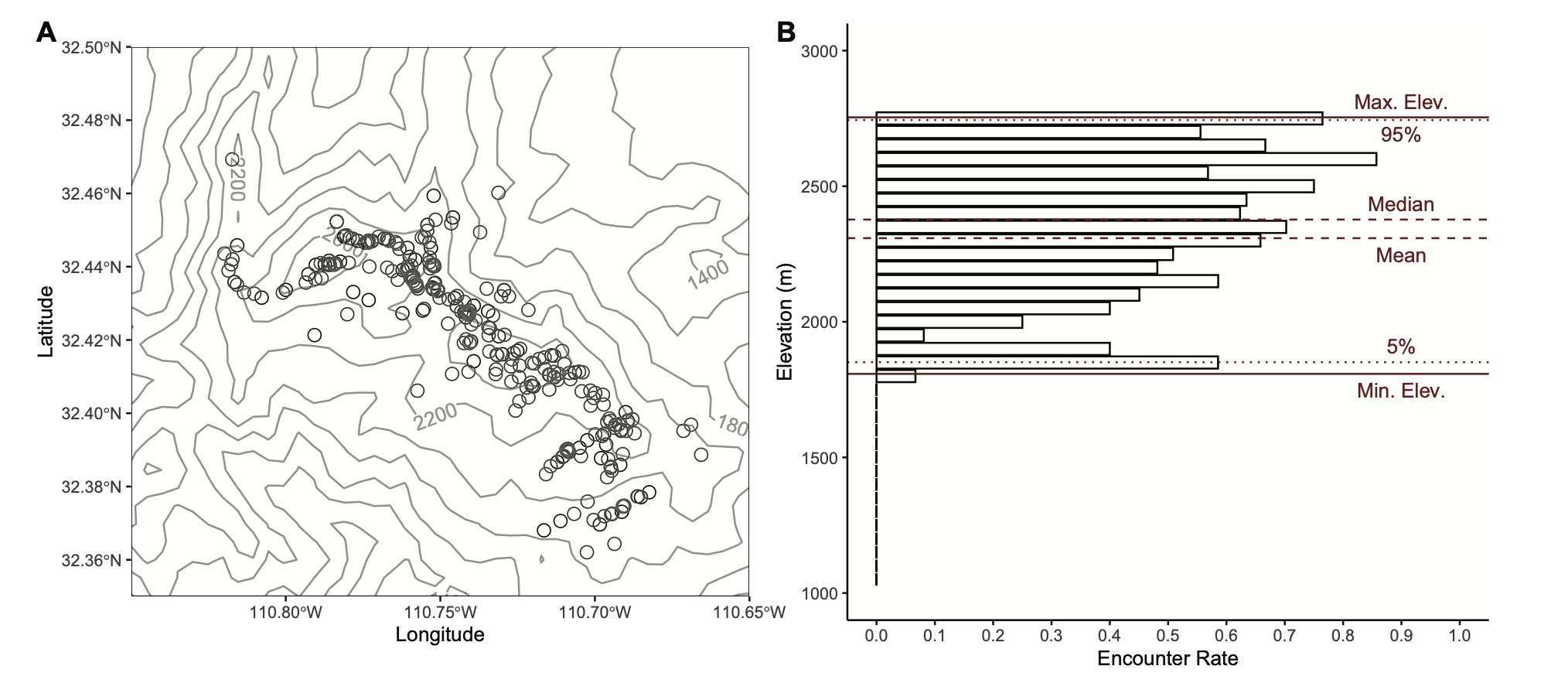
- Conservation genomics and climate adaptation in American Pika at their southern range limit (led by Ph.D. student Elizabeth Osterhoudt; funded by the Western National Parks Association)
Hybridization and Speciation
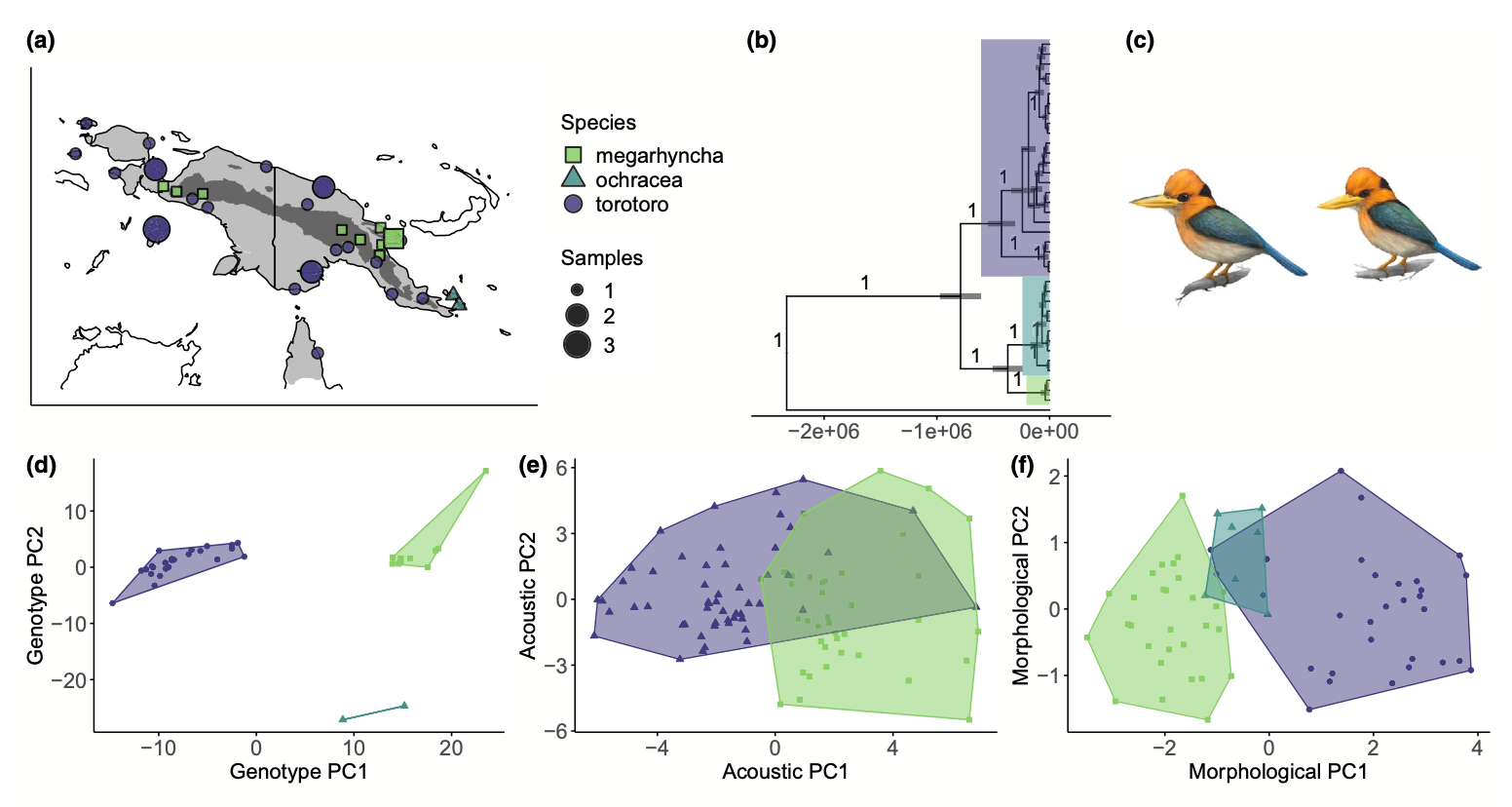
The widespread application of high throughput sequencing technology to nonmodel organisms beginning in the early 2010s revealed that gene flow between species was more pervasive than previously thought, casting traditional models of speciation in strict geographic isolation into question. The lab has longstanding interest in using genomics, simulations and modeling, and field observations to identify the ecological and evolutionary processes that permit divergence and the development of reproductive isolation despite ongoing or intermittent interbreeding. Previous work in this area has included a study of speciation and gene flow between elevationally segregating sister species (Linck et al. 2020; with Ben Freeman and Jack Dumbacher) and the genetic basis of reproductive isolation in passerine hybrid zones (Semenov et al. 2021 and Semenov et al. 2025; with Georgy Semenov, Scott Taylor, Matt Carling, Zac Cheviron, Amber Rice, and others).
Current projects:
- Gene flow and ecotypic divergence in Melanesian parrotfinches (with Lucas DeCicco, Devon DeRaad, Brett Benz, and Rob Moyle)
Evolutionary Ecology of Elevational Ranges
Plants and animals typically have constrained elevational distributions, leading to rapid turnover in species and communities from the base of mountains to their summit. This pattern provides an opportunity to disentangle the factors that limit species ranges more from the confounding effect of geographic distance. Because many species worldwide are moving upslope in response to climate change, elevational ranges have also become a subject of intense interest in global change biology. PI Linck has been fascinated by the elevational distributions of organisms since seeing his first Bicknell’s Thrush near the summit of Mt. Ellen in Vermont in high school. Previous work in this area has used evolutionary history to infer climate vulnerability in tropical birds (Linck et al. 2021; with Ben Freeman, Daniel Cadena, and Cam Ghalambor), explored the relationship between blood traits related to hypoxia tolerance and elevational range characteristics (Linck et al. 2023 and Williamson et al. 2023; with Chris Witt and Jessie Williamson), and demonstrated lowland species are shifting their ranges most rapidly (Mamantov et al. 2020; with Maggie Mamantov, Dan Gibson-Reinemer, and Kimberly Sheldon). PI Linck also recent published synthesis of conceptual issues related to elevational ranges (Linck 2025).
Current projects:
- A comparison of point count data and eBird records for describing elevational variation in abundance (led by undegraduate Nate Ingelbinger)
- Molecular convergence in hemoglobin subunits in high elevation Diglossine tanagers (with Chris Witt and others)
Natural History and Ornithology in Melanesia
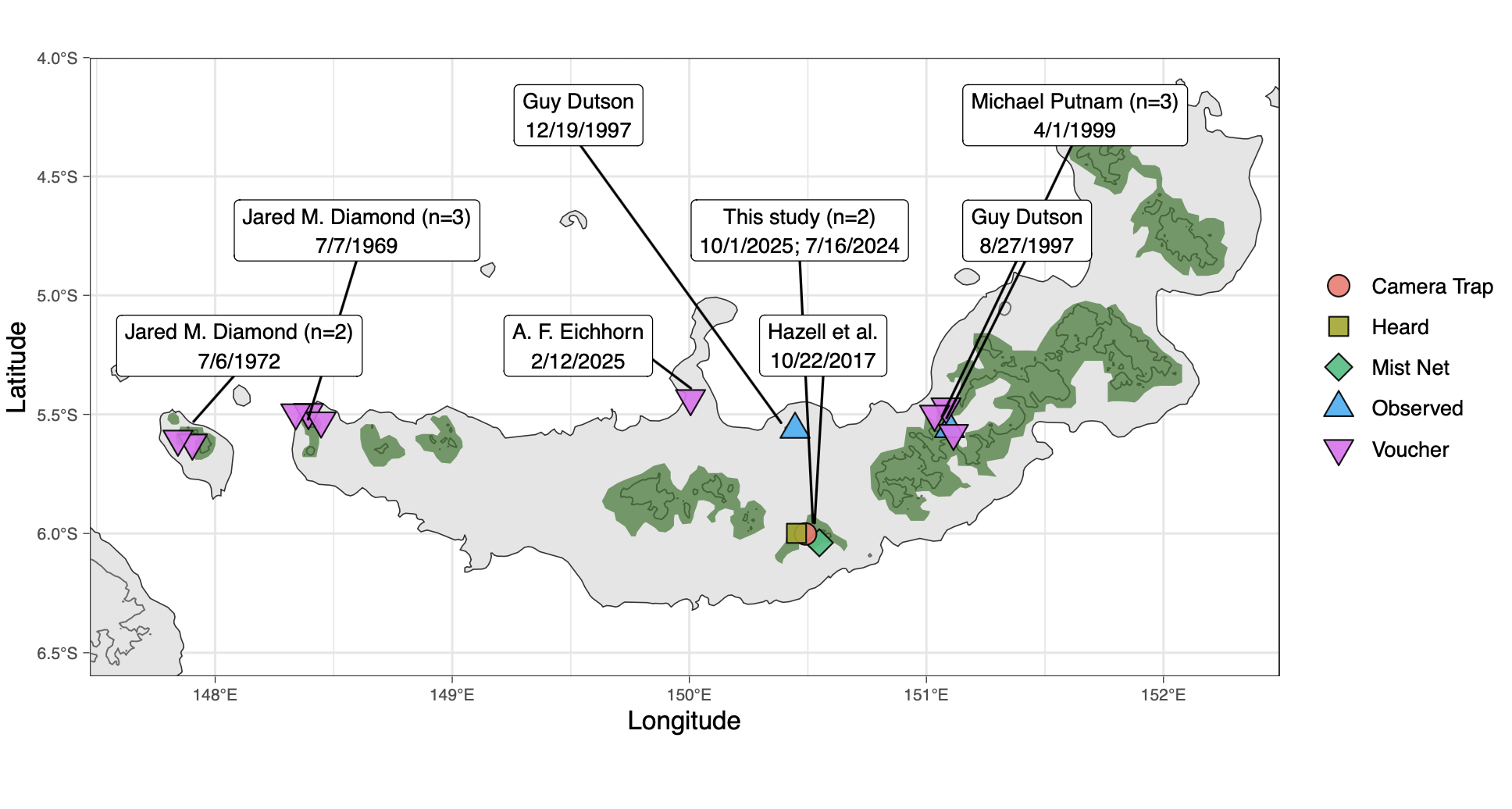
PI Linck has a long history of traveling and working in Papua New Guinea and the southwest Pacific, a region with high biodiversity wilderness areas, imperiled biodiversity hotspots, and strong Indigenous land tenure. Despite their historical importance to the development of ecology and evolutionary biology through seminal work by Alfred Russel Wallace, Ernst Mayr, Jared Diamond, and others, birds in this region remain poorly known. In addition to contributing samples and specimens to natural history museum collections to support evolutionary biology research—some of the only modern material from the region—we conduct rapid assessments of avian diversity in regions that have rarely or never been visited by formally trained ornithologists. In this way we have documented two species formerly lost to science since 2024: the New Britain Thrush and the New Britain Bronzewing.
Current projects:
- Avian biodiversity surveys in West New Britain (with M.S. student Jason Gregg, Bulisa Iova, and Joe Wood
- The impact of oil palm expansion on Local Ecological Knowledge (with M.S. student Jason Gregg and Alfred Kik)